|
Complement
COMPONENTS AND FUNCTIONS OF THE COMPLEMENT SYSTEM
The complement system found in the blood of mammals is composed of heat labile substances (proteins) that combine with antibodies or cell surfaces. This complex, multicomponent system is composed of about 26 proteins. The "complement cascade" is constitutive and non-specific but it must be activated in order to function. The functions of complement include:
- making bacteria more susceptible to phagocytosis
- directly lysing some bacteria and foreign cells
- producing chemotactic substances
- increasing vascular permeability
- causing smooth muscle contraction
promoting mast cell degranulation
The complement system can be activated via two distinct pathways; the classical pathway and the alternate pathway. Once initiated, a cascade of events (the "complement cascade") ensues, providing the functions listed above.
Most of the complement components are numbered (e.g. C1, C2, C3, etc.) but some are simply refered to as "Factors". Some of the components must be enzymatically cleaved to activate their function; others simply combine to form complexes that are active. The following table lists these components and their functions.
| Components of the Classical Pathway |
| Native component |
Active component(s) |
Function(s) |
| C1(q,r,s) |
C1q |
Binds to antibody that has bound antigen, activates C1r. |
| C1r |
Cleaves C1s to activate protease function. |
| C1s |
Cleaves C2 and C4. |
| C2 |
C2a |
Unknown. |
| C2b |
Active enzyme of classical pathway; cleaves C3 and C5. |
| C3 |
C3a |
Mediates inflammation; anaphylatoxin. |
| C3b |
Binds C5 for cleavage by C2b.
Binds cell surfaces for opsonization and activation of alternate pathway. |
| C4 |
C4a |
Mediates inflammation. |
| C4b |
Binds C2 for cleavage by C1s. Binds cell surfaces for opsonization. |
| Components of the Alternate Pathway |
| Native component |
Active component(s) |
Function(s) |
| C3 |
C3a |
Mediates inflammation; anaphylatoxin. |
| C3b |
Binds cell surfaces for opsonization and activation of alternate pathway. |
| Factor B |
B |
Binds membrane bound C3b. Cleaved by Factor D. |
| Ba |
Unknown. |
| Bb |
Cleaved form stabilized by P produces C3 convertase. |
| Factor D |
D |
Cleaves Factor B when bound to C3b. |
| Properdin |
P |
Binds and stabilizes membrane bound C3bBb. |
| Components of the Membrane-Attack Complex |
| Native component |
Active component(s) |
Function(s) |
| C5 |
C5a |
Mediates inflammation; anaphylatoxin, chemotaxin. |
| C5b |
Initiates assembly of the membrane-attack complex (MAC). |
| C6 |
C6 |
Binds C5b, forms acceptor for C7. |
| C7 |
C7 |
Binds C5b6, inserts into membrane, forms acceptor for C8. |
| C8 |
C8 |
Binds C5b67, initiates C9 polymerization. |
| C9 |
C9n |
Polymerizes around C5b678 to form channel that causes cell lysis. |
ACTIVATION OF THE COMPLEMENT CASCADE
Classical Pathway
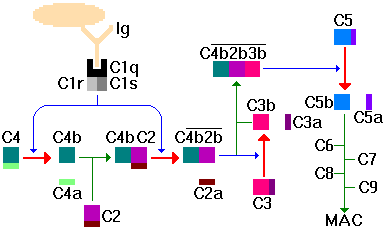
The classical pathway starts with C1; C1 binds to immunoglobulin Fc (primarily IgM and IgG); C1 is recognition complex composed of 22 polypeptide chains in 3 subunits; C1q, C1r, C1s. C1q is the actual recognition portion, a glycoprotein containing hydroxyproline and hydroxylysine that looks like a tulip flower. Upon binding via C1q, C1r is activated to become a protease that cleaves C1s to a form that activates (cleaves) both C2 and C4 to C2a/b and C4a/b. C2b and C4b combine to produce C3 convertase (C3 activating enzyme). C4a has anaphylactic activity (inflammatory response).
C3 is central to both the classical and alternative pathways. In classical, C4b2b convertase cleaves C3 into C3a/b. C3a is a potent anaphylatoxin. C3b combines with C4b2b to form C4b2b3b complex that is a C5 convertase. C3b can also bind directly to cells making them susceptible to phagocytosis.
C5 is converted by C5 convertase (i.e. C4b2b3b) to C5a/b. C5a has potent anaphylatoxic and chemotaxic activities. C5b functions as an anchor on the target cell surface to which the lytic membrane-attack complex (MAC) forms. MAC
includes C5b, C6, C7, C8 and C9. Once C9 polymerizes to form a hole in the cell wall, lysis ensues.
Alternate Pathway
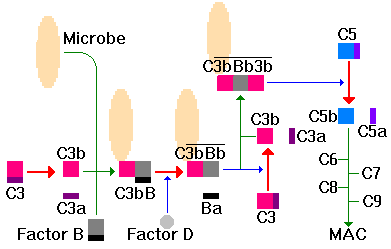
The alternate pathway may be initiated by immunologic (e.g. IgA or IgE) or non-immunologic (e.g. LPS) means. The cascade begins with C3. A small amount of C3b is always found in circulation as a result of spontaneous cleavage of C3 but the concentrations are generally kept very low (see below). However, when C3b binds covalently to sugars on a cell surface, it can become protected. Then Factor B binds to C3b. In the presence of Factor D, bound Factor B is cleaved to Ba and Bb; Bb contains the active site for a C3 convertase. Next. properdin binds to C3bBb to stabilize the C3bBb convertase on cell surface leading to cleavage of C3. Finally, a C3bBb3b complex forms and this is a C5 convertase, cleaving C5 to C5a/b. Once formed, C5b initiates formation of the membrane attack complex as described above.
Generally, only Gram-negative cells can be directly lysed by antibody plus complement; Gram-positive cells are mostly resistant. However, phagocytosis is greatly enhanced by C3b binding (phagocytes have C3b receptors on their surface) and antibody is not always required. In addition, complement can neutralize virus particles either by direct lysis or by preventing viral penetration of host cells.
REGULATION OF THE COMPLEMENT CASCADE
Because both the classical and alternate pathways depend upon C3b, regulation of the complement cascade is mediated via 3 proteins that affect the levels and activities of this component.
- C1 Inhibitor inhibits the production of C3b by combining with and inactivating C1r and C1s. This prevents formation of the C3 convertase, C4b2b.
- Protein H inhibits the production of C3b by inhibiting the binding of Factor B to membrane-bound C3b, thereby preventing cleavage of B to Bb and production of the C3 convertase, C3bBb.
- Factor I inhibits the production of C3b by cleaving C3b into C3c and C3d, which are inactive. Factor I only works on cell membrane bound C3b, mostly on red blood cells (i.e. non-activator surfaces).
|
|